Overview
This
lecture focusses on the idea that some part(s) of the brain control
feeding. As you read this material you may form the impression that
things have not gone very well in this area over the years. To some
extent this is true. A lot of the early research was influenced by
theoretical models of motivation that were popular at the time. A
considerable amount of effort was devoted to discovering areas of the
brain that controlled motivation. You may consider that things were not
helped by the early discovery of discrete nuclei in the brain which -
when lesioned - had the dramatic effect of either increasing or
decreasing body weight and food intake. These results were seized upon
as being possible physiological substrates for the psychological states
of hunger and satiety. In fact subsequent research has revealed that
lesions can have more than one effect, and that the brain consists of
circuits affecting behaviour(s). This lecture does not attempt to give
an exhaustive coverage of the area. Instead we will look at two brain
areas implicated in the control of eating, and see how ideas about
their roles have changed over the last half century.
Do not
panic, your computer screen is not on the blink! The picture of the
Suomo at the top of the page is a negative to reflect the focus of this
lecture on things that happen beneath the skin.
The
Dual-Control Theory of Feeding
|
Dual-control
theory was based on a homeostatic
view of hunger and satiety.
- A
decline in glucose activated the lateral hypothalamus (LH)
- Activity
within the LH gave rise to hunger
- Hunger
motivated the search for and consumption of food
- Food
was broken down to release glucose
- Glucose
activated the ventromedial hypothalamus (VMH)
- Activation
of the VMH caused a feeling of satiety
- Satiety
inhibited further feeding
|
|
The
diagrams below show the location of the LH and VMH, and a diagrammatic
representation of the relationship between physiology, motivation and
brain nuclei.
| Point to ponder: Study
the diagram and see if you can see why the system - as set out in the
diagram - might not maintain homeostasis. |
 |
Cross
section of rat brain showing approximate location of LH and VMH

|
Evidence
for Control Centres for Feeding.
The
idea that the VMH acts as the brain's satiety centre
whereas the LH is a hunger centre, was put forward
by Eliott Stellar in 1954 (Psychological Review, 61, 5-22) and it
dominated thinking about feeding behaviour for the next 20 years. It
was supported by the following set of experimental findings on the
effects of removing, or electrically activating each area
| Area of hypothalamus
|
Effect of lesioning
|
Effect of stimulating
|
| Ventromedial
hypothalamus |
Increases
eating |
Decreases
eating |
| Lateral hypothalamus |
Decreases
eating |
Increases
eating |
Lesions to the VMH and LH produced the most straightforward and
startling effects. This animation should help you visualize the
location of the centres within the brain. As you examine the path taken
by the lesioning electrodes consider how much 'collateral damage' is
involved. How you would design an experiment to rule out the
possibility that the effects of the lesion were in fact due to
'collateral damage' rather than destruction of a particular
hypothalamic nuclei?
| Lesion of
ventromedial hypothalamus (VMH) |
Lesion of
lateral hypothalamus (LH) |
|
|

|

|
|
Effect
of VMH Lesions on Eating
 |
Compared
to controls VMH lesioned rats show:
- excessive
eating ( hyperphagia )
- weight
gain
The
effects of the lesion are not permanent. After an initial increase,
body weight stabilizes.
To
reflect this VMH lesioned rats are said to be in either a
- dynamic
or
- static
phase of weight gain
It
is clear that the simple dual point theory cannot explain the
results.
VMH
rats are able to reach satiety despite the absence of their satiety
centre.
|
Effect
of VMH Lesions on Hunger
|
The
dual centre theory predicts that lesioning the VMH will increase:
- consummatory
behaviour (eating)
- hunger
(motivation to eat)
Although
the consummatory behaviour of rats with
VMH lesions is consistent with the theory, the motivational
effects of VMH lesions are less clear-cut. This diagram shows the
willingness of rats to press a lever to obtain food as the demands of
the schedule are increased from a starting point where each bar press
is rewarded with food (FR1), right up to a schedule where the rat must
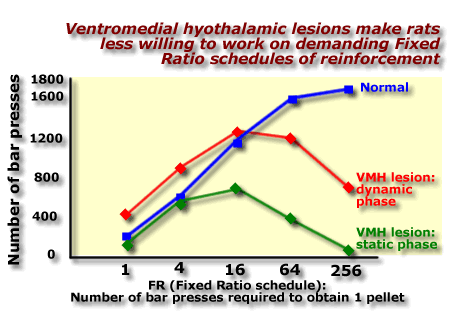
press 256 times to get one pellet (FR256). [Source: Teitelbaum, 1957]
The
logic behind this experiment is "If you are hungry, you will be
willing to work hard to get food". The results show that intact rats
press the bar more as the schedule becomes more demanding. VMH lesioned
rats in the dynamic phase press more frequently than normal rats when
the less demanding schedules (FR1 through FR16) are in operation, but
are less willing to work under the more demanding conditions imposed by
FR64 and FR256 schedules of reinforcement. This laziness
is even greater in VMH lesioned rats who are in the static phase.
There
is a body of evidence which indicates that VMH lesioned rats are
finicky eaters. Compared to normal rats they eat less when:
- quinine
is added to make food taste bitter
- food
is stale
These
are not the behaviours you might expect if the VMH is a hunger
centre whose destruction increases an animal's motivation for food!
These
animal experiments appear to correspond to the observation that -
compared to normal weight people - obese humans
eat fewer peanuts if they have to shell them.
|
Effect
of work on the eating behaviour of normal and obese humans
| |
|
Number
who |
| Eat
|
Don't eat
|
| Normal subjects
|
Nuts have
Shells |
10 |
10 |
| Nuts have
no shells |
11 |
9 |
| Obese subjects |
Nuts have
Shells |
1 |
19 |
| Nuts have
no shells |
19 |
1 |
|
Effect
of LH Lesions on Eating
|
The
diagram below shows that initially, animals with lateral hypothalamic
lesions exhibit severe loss of body weight due to their refusal to eat
( aphagia ) and drink ( adipsia
). But these effects are not permanent. Body
weight does not decline inexorably. The table shows that rats go
through 4 stages of recovery after LH lesions.
Dual point theory cannot explain these results.
|
|

|
LH
lesioned rats recover the ability to eat despite the absence of their
feeding centre.

|
Set
Point Theory
|
We
have seen that the Dual-Centre hypothesis - which visualized the LH as
a feeding centre and the VMH as a satiety centre - could not explain
the long term effects of lesions to these areas. One solution to this
problem was to suggest that these nuclei controlled body weight through
a set-point mechanism .
According
to this view:
- LH
lesions reduced the
set
point for
body weight. Lesioned rats maintain body weight at a new
lower level
- VMH
lesions increased the
set
point
for body weight . Lesioned rats maintain body weight at a new
higher level
|
|
 One
way of testing this idea was to starve rats so that they lost body
weight before receiving LH lesions. According to
set-point theory LH lesions should have little or no effect on these
rats because starvation would reduce body weight so that it was already
at the new set-point before the operation. The diagram below shows that
preoperative starvation did have the effect predicted by set-point
theory. LH lesions did not cause a further reduction in body weight. One
way of testing this idea was to starve rats so that they lost body
weight before receiving LH lesions. According to
set-point theory LH lesions should have little or no effect on these
rats because starvation would reduce body weight so that it was already
at the new set-point before the operation. The diagram below shows that
preoperative starvation did have the effect predicted by set-point
theory. LH lesions did not cause a further reduction in body weight.
Other
experiments
on rats made obese by VMH lesions revealed that
if they were force fed (so that their body
weight was increased ) and then allowed to feed ad lib, they would
loose weight until they returned to the weight they reached during the
static phase.
This
result supports the hypothesis that VMH lesions increased the set
point for body weight
|
|
LH
Lesions Cause Sensory Neglect
Marshall et al (1971) found that LH lesions produce a range of sensory
and motor impairments termed sensory neglect .
Rats
with LH lesions
- Do
not respond to touch, auditory, or visual stimuli
- Remain
immobile unless disturbed
- Do
not right themselves when placed on their side
This
experiment involved a very elegant design to ensure that the observed
effects were not due to some non-specific effect such as illness.
It is
clear from the diagram
that there are two lateral hypothalamic nuclei; one on each side of the
brain.
Marshall lesioned the LH on one side of the brain (called a
unilateral lesion) and left the LH on the other side of the
brain intact. He found that sensory neglect was only shown when one
side of the body was stimulated. If the same stimulus was presented to
the other side of the body, the rat reacted normally. This shows that
the rat was able to respond. It was not
suffereing from general malaise.
(Source: Marshall et al, Science, 174, 523-525, 1971)
Nigrostriatal
Lesions Produce Aphagia and Adipsia
|
|
One implication
of Marshall et al's findings is that LH lesions may reduce
arousal . You may recall Positive Incentive
Theory (described in an earlier lecture), which suggests
that eating is triggered by external stimuli
such as the time of day, or the sight and smell of food. If LH lesions
interfere with an animal's ability to process stimuli this could
account for reduced food intake. Obviously this explanation casts doubt
on the idea that the LH is specifically involved in food intake; all
behaviour would be affected if sensation was disrupted. It is now
thought that the sensory neglect produced by LH lesions is caused by
the lesion destroying axons in the nigrostriatal
tract that pass through the lateral hypothalamus
and terminate in the caudate-putamen.
The
animation shows the impact of LH lesions on the nigrostriatal
pathway.
The
animation shows how lesions - aimed at the LH - would damage axons of
the nigrostriatal system at the point where they
pass through the lateral hypothalamus. You may remember that we studied
this system in a previous lecture. The nigrostriatal tract utilizes
dopamine ( DA ) as its transmitter.
Parkinson's disease is associated with a massive reduction
in dopamine in the caudate nucleus.
|
One
implication
of Marshall et al's findings is that LH lesions may reduce
arousal . You may recall Positive Incentive
Theory (described in an earlier lecture), which suggests
that eating is triggered by external stimuli
such as the time of day, or the sight and smell of food. If LH lesions
interfere with an animal's ability to process stimuli this could
account for reduced food intake. Obviously this explanation casts doubt
on the idea that the LH is specifically involved in food intake; all
behaviour would be affected if sensation was disrupted. It is now
thought that the sensory neglect produced by LH lesions is caused by
the lesion destroying axons in the nigrostriatal
tract that pass through the lateral hypothalamus
and terminate in the caudate-putamen.
The
animation shows how lesions - aimed at the LH - would damage axons of
the nigrostriatal system at the point where they
pass through the lateral hypothalamus. You may remember that we studied
this system in a previous lecture. The nigrostriatal tract utilizes
dopamine ( DA ) as its transmitter.
Parkinson's disease is associated with a massive reduction
in dopamine in the caudate nucleus.
|
Ungerstedt
(Acta Physiol. Scand. 82, (Suppl. 367, 69-93, 1971) carried out a
series of elegant experiments that show that the sensory
neglect following LH lesions is probably the result of
collateral damage to the nigrostriatal
DA system.
Ungerstedt
injected the neurotoxin (chemical nerve poison)
6-hydroxydopamine [6-OHDA] into the cell bodies of the
nigrostriatal tract which are found in the substantia nigra
. The animated diagram illustrates the important point that the
substantia nigra lies caudal to the LH, so any effect of the chemical
lesion cannot be attributed to LH damage. A
separate group of rats were given conventional (electrical ) lesions in
the LH.
Results
: Both types of lesion (LH and substantia nigra)
produced the "lateral hypothalamic syndrome" i.e. initial apahagia and
adipsia followed by a gradual but complete recovery of food and water
intake.
|
|
Chemical
Lesions of the LH
We
have seen that electrical lesions of the LH have failed to provide
clear cut answers to the role of the LH in feeding. Chemical lesions on
the other hand did provide a resolution of the issue of sensory
neglect. Recently research has employed chemical lesions to destroy
cells in the LH whilst sparing axons of the
nearby nigrostriatal tract. Kainic acid and
ibotenic acid lesions of cells in the LH produce a
long-lasting decrease in food intake and body weight, but they do not
produce sensory neglect (Winn et al, Neuroscience, 12, 225-240, 1984).
However it is widely acknowledge that it is a gross oversimplification
to call the LH a 'feeding centre' because:
- Animals
recover
from LH lesions
- LH
lesions disrupt aggression, sexual behaviour, and reinforcement
- Nigrostriatal lesions
produce aphagia and adipsia
Role
of the LH & VMH - What do contemporary textbooks say?
The
search for a role for the LH in hunger
continues. It may play a role in controlling insulin release. According
to this idea, excitation in the LH stimulates the release of insulin
from the pancreas. Lesioning the LH is followed by a reduction in
insulin release. Insulin allows glucose to be broken down to provide a
source of energy. The body compensates for the lack of glucose-derived
energy by breaking down stored body-fat to provide fuel. As a result of
fat-breakdown, body weight declines. But the blood is full of excess
energy-rich free fatty acids and the animal fails to eat because its
body is telling the brain areas controlling feeding that there is no
need to take in more energy (Schneider & Tarshis, Elements of
Physiological Psychology, McGraw Hill, 1995, p 386-7, 1995).
Pinel
( Biopsychology , Allyn & Bacon,p260-261, 2000) has provided a
complementary explanation for the role of the VMH
in feeding. He suggests that animals with VMH lesions "
overeat because they become obese ". He argues that VMH
lesions increase blood insulin levels. This triggers lipogenesis (the
conversion of glucose into fat). Because any food the animal eats is
rapidly converted into fat, they suffer an energy deficiency.
Consequently they eat in a vain attempt to overcome the lack of glucose
circulating in their bloodstream.
Copyright Dr.
C.A.P. Kenyon 1994-2006

