|
|
 |
Visit the SALMON Bookshop for recommended books on this topic |
|
|
|
Visit the SALMON Bookshop for recommended books on this topic |
The aim of this lecture is to show you how careful experimental investigation is needed to test even the simplest theory. You will be presented with a series of experiments that suggest that although some aspects of the effect of food deprivation on eating are consistent with drive reduction theory, other factors such as variations in eating as a function of time of day, meal size, and the spacing of meals require a more sophisticated interpretation.
It is important to bear in mind that because an experiment contains a design fault, or a theory does not withstand scrutiny, this does not mean that the effort that went into the theory or experiment was wasted. On the contrary, a theory that appears weak with the benefit of hindsight was useful if it generated experiments that led to greater theoretical insights. Likewise an experiment that appears flawed to our eyes, may have been conducted at a time when psychological research methodology was in its infancy. Our present day sophistication is built on a raft of early experiments that may appear naive to our eyes. But we should remember that future generations will look back on our efforts with a knowing smile.
The lectures ends with a discussion of early physiological theories that stressed the importance of maintaining a balance of glucose and fat reserves within the body. These physiological notions are very similar to the drive reduction theory of eating behaviour. This is not surprising because early physiological and behavioural theories were based on the idea of the body striving to maintain a homeostatic balance. A modern reinterpretation of glucostatic theory is presented which mirrors contemporary psychological theories which stress the importance of incentive stimuli creating motivation
The term motivation is used to describe a force that impels us to behave in particular ways. This lecture will draw upon two types of motivational theory:
|
According to drive reduction theory food deprivation creates
|
Operation
of drive
reduction theory:
|
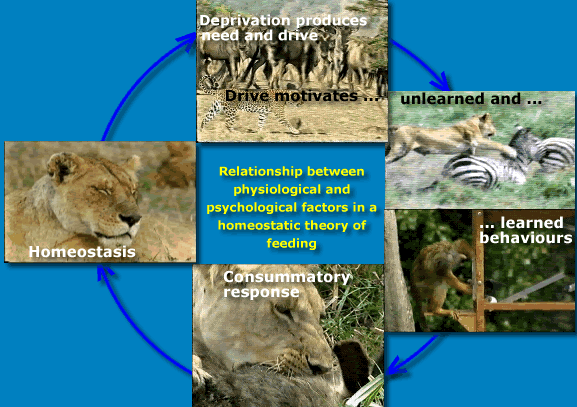 |
This view of hunger has its roots in Cannon's concept of homeostasis which he developed to describe dynamic equilibrium in physiological systems. If part of an animal is disturbed (e.g. if body tissue is not receiving enough oxygen or food) the animal behaves in a way that will alleviate the deficiency, and restore the homeostatic balance. According to this theory, the hunger drive serves to maintain the body's energy balance. In other words, hunger is due to deprivation, and the animal's behaviour is designed to reduce deprivation. |
Internal hypothetical states - such as drive - are hidden within the animal and therefore impossible to observe and measure. The amount of internal drive has to be inferred from the animals external observable behaviour.
 |
We
can examine the relationship between observable factors
such as:
It is difficult and laborious to accurately measure biting, chewing and swallowing food. It is much easier to measure the amount of food consumed, or the number of bar presses emitted in order to obtain a predetermined amount of food. A simple homeostatic model predicts that food intake will increase linearly (up to some theoretical maximum) as a function of the duration of preceding deprivation. In other words, the longer an animal had been without food, the more it will eat when finally given access to food. |
However, reality is more complicated than this simple prediction. In fact, as the next experiment reveals, food intake appears to be a function when the animal is allowed to eat, as well as how long it has gone without food
Bare (1959) trained rats
to bar press for food, and then allocated them to Deprived or
Control groups.
|
 |
Laboratory rats are nocturnal - they are more active during the night than during human daytime.
Bare's results show that:
Conclusion: The amount of food eaten by food deprived rats is a function of food deprivation, and the time of day they are allowed to eat.
Source: Bare ( Journal of Comparative and Physiological Psychology , 52, 129-131,1959)
A weakness of this experiment is that food deprivation is confounded with time of testing. In other words there are two things that vary between the different food deprived groups:
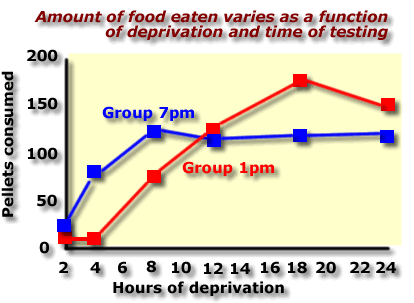
In his next experiment Bare held the time of testing constant and varied only one factor: duration of deprivation
Bare
& Cicala (1960) varied the duration of deprivation between 2
and 24 hours for separate groups of rats and tested independent groups
at 1pm and 7pm
The results show that eating appears be influenced by the time of testing as well as the length of deprivation.
Source: Bare & Cicala, Journal of Comparative and Physiological Psychology ,53, 151-154, 1960 |
 |
||||||||||||||||||||||||
Another important variable controlling the intake of deprived rats is the rats experience of deprivation, specifically the number of days the animal has been on a particular deprivation schedule.
| Reid & Finger
(1955) maintained a group of rats on a 23 hour deprivation schedule
which only allowed access to food for one hour each day.
They found that these rats stabilized their daily intake after they had been exposed to the schedule for over 20 days. It is also clear that the amount eaten during the single one-hour mealtime never reached the amount consumed by rats that were allowed access to food throughout the day. Clearly these results are inconsistent with a simple homeostatic model of food intake. Source: Reid & Finger ( Journal of Comparative and Physiological Psychology , 48, 10-113, 1955) |
 |
 The
homeostatic account of eating has
also been challenged on the grounds that it does not reflect food
regulation as it occurs
under natural conditions. Guinea pigs fed ad lib (food available
throughout the day)
always have food in their stomachs. This indicates that there is always
a reservoir of
food in the stomach that can be absorbed, and that the animals does not
experience food
deprivation.
The
homeostatic account of eating has
also been challenged on the grounds that it does not reflect food
regulation as it occurs
under natural conditions. Guinea pigs fed ad lib (food available
throughout the day)
always have food in their stomachs. This indicates that there is always
a reservoir of
food in the stomach that can be absorbed, and that the animals does not
experience food
deprivation.
This cheetah has a distended stomach after gorging itself on prey. Observation of animals in the wild suggests that they take the opportunity to eat whenever possible.
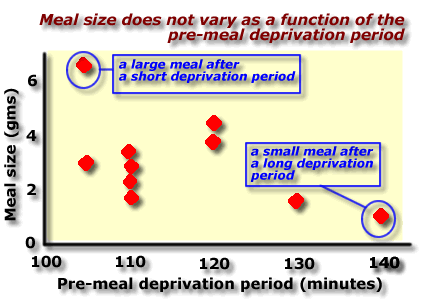
| Le Magnen (1971)
examined meal sizes in rats maintained on an ad lib
food schedule.
He found that rats ate their daily intake in 8 to 10 meals of variable size (averaging 2 gms), separated by periods of no eating of various lengths. Le Magnen went on to investigate the relationship between meal size and meal-to-meal interval. According to homeostatic theory, meal size should increase with increasing time since the last meal (i.e. the period of deprivation). In fact these results show that the amount eaten in a meal bears no relation to the length of time since the last meal. Source: Le Magnen ( Progress in Physiological Psychology , Volume 4, 1971) |
 |
However Le Magnen did find a highly significant correlation between meal size and the amount of time the animal would wait before eating again.
| If the rat eats a large
meal it will wait a longer time before eating again, so the onset
of eating appears to be dependant on need.
Le Magnen's findings suggest that the amount of eaten during a meal is independent of physiological need, perhaps other factors such as the taste of food, or the amount of food already in the rat's stomach determine how much will be eaten. Source: Le Magnen ( Progress in Physiological Psychology , Volume 4, 1971) This effect is not restricted to rats. De Castro and De Castro (1989) have reported a correlation in humans between the time since the previous meal, and the size of the present meal, but only when people eat alone. The correlation disappears when people eat in groups. Source: De Castro and De Castro ( American Journal of Clinical Nutrition , 50, 237-247, 1989) |
 |
|
|
According
to positive incentive theory we eat
three meals a day out of habit,
or because of external stimuli such as the time of
day, or the sight and smell of
food.
Demonstrations of classically conditioned eating in humans and animals are consistent with this idea
|
Classical Conditioning of Food Intake.
| Weingarten (19830
demonstrated classical conditioning of eating in rats.
Training procedure: Hungry rats were presented with
Testing procedure : Rats given ad lib access to food
Results :
Conclusion
Source: Weingarten( Science , 220, 431-432, 1983) |
 |
|
|
Everyone is aware of
their ability to eat more if the meal is composed of a variety of
foods. For example, we often consume a tasty pudding after a main
course of meat and potatoes, even though we could not eat any more meat
and potatoes. Sensory-specific satiety (SSS)
refers to the decrease in the pleasantness and consumption of food
after eating it to satiety.
Much of the work in this area has been carried out by the Rolls at Oxford. They find that it is not just the taste of food that influences SSS, colour and shape also influence consumption. In the results illustrated here, consumption of sandwiches was affected by the variety of fillings:
These results are consistent with the positive incentive theory, and would be difficult to explain in terms of drive reduction. |
|

|
 |
|
| Source: Rolls, Rolls
& Rowe, Physiology and Behaviour , 29, 409-417, 1982
I have been influenced by the principle of sensory-specific satiety in preparing these diagrams. I reckon the addition of pictures as backgrounds to the diagrams may overcome the satiety you may have built up after exposure to a series of bland diagrams! |
||
 In the
1940s and 1950s, experimental research was interpreted as supporting a
fairly simple relationship between physiology, hunger and body weight. In the
1940s and 1950s, experimental research was interpreted as supporting a
fairly simple relationship between physiology, hunger and body weight.
|
 |
 |
A considerable body of
experimental evidence can be interpreted in terms of Glucostatic
Theory .
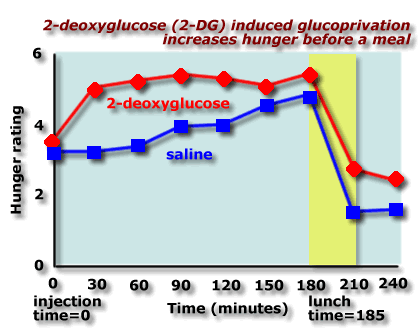
For example, a chemical called 2-deoxyglucose (2-DG) competes with glucose in the body:
These results show that people given 2-DG report more hunger than those given saline. Source: Thompson & Campbell, Science , 198, 1065-1068, 1977 |
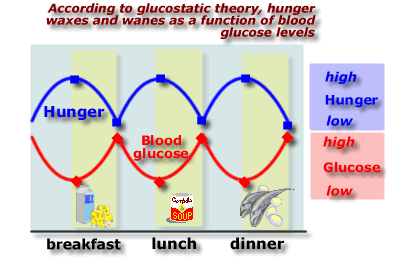
According to Glucostatic Theory, as the body consumes energy in the form of glucose, glucose level in the blood declines and this initiates feeding to maintain homeostatic energy balance. The next experiment appears to provide support for this view, but there is a provocative alternative explanation for these results.
| Campbell and Smith
(International Journal of Obesity, 14 (supplement 3), 15-33,1990) have
reported a transient decline in blood glucose shortly before rats began
their next meal. These results could be used to support the Glucostatic
Theory. However, not all agree with this interpretation. Pinel (
Biopsychology , Allyn & Bacon,p 259,2000 ) argues that the
decline in glucose is preceded by the release of insulin which is
triggered by the animals intention to eat. He
suggests that the suddenness of the drop in glucose indicates that the
rats lowered their own glucose by releasing insulin, rather than being
a consequence of a gradual decline in energy reserves.
This provocative idea is consistent with a Positive Incentive Theory of Eating but would not be predicted by Homeostatic Theories (e.g. drive reduction and glucostatic theories). Pinel's views are in line with an idea put forward by Woods ( Psychological Review , 98, 488-505, 1991). Woods constructs an explanation for hunger that is exactly the opposite of a conventional homeostatic theory of eating. He argues that
|
 |
Logue,A.W.
(1998). Evolutionary
Theory and the
Psychology of Eating. Logue's paper discusses how
evolutionary theory can be used as a
theoretical framework to help psychologists understand one particular
type of behavior:
the consumption of food. Here are some relevant highlights from this
useful article:
Abed (1998) The sexual competition hypothesis for eating disorders. British Journal of Medical Psychology 71(4):525-547.
Abstract. A hypothesis is presented for eating disorders, based on Darwinian theory, that contends that these syndromes together with the phenomenon of the pursuit of thinness are manifestations of female intrasexual competition. It is suggested that eating disorders originate in the human female’s psychological adaptation of concern about physical attractiveness which is an important component of female ‘mate attraction’ and ‘mate retention’ strategies. It is argued that present day environment of Western countries presents a range of conditions which have led to the over-activation or the disruption of the archaic female sexual strategy of maximising ‘mate value’. The present hypothesis deals with the ultimate level of causation and is therefore compatible with a range of theories of proximate causation. Although the present hypothesis is not directly testable, it makes predictions that are testable and refutable. Finally it is suggested that the sexual competition hypothesis has more explanatory power than existing evolutionary theories of eating disorders.

|
A hormone called ghrelin stimulates feeding in rodents.The concentration of ghrelin in a person's blood rises significantly before meals and plummets afterwards. A recent study reports that people receiving premeal infusions of the hormone increased their food intake by an average of 30 percent. more ... |
Global obesity: In 1995, there were an estimated 200 million obese adults worldwide. Another 18 million children under-five were classified as overweight. By 2000, the number of obese adults had increased to more than 300 million and in developing countries it is now estimated that more than 115 million people suffer from obesity-related problems. more ...
Copyright Dr. C.A.P. Kenyon 1994-2006



