
|
Visit the SALMON Bookshop for recommended
books on this topic
|
|
|
|
Visit the SALMON Bookshop for recommended
books on this topic
|
|
Overview:
New-born rat pups appear helpless little pink bundles of potential
behaviour. Don't be
fooled, they are already behaving their little heads off! You may think
they are
"helpless passive recipients of maternal care", but they are from it.
They are
engaged in an intricate behavioural dance with their parents for
survival. New-born pups
play an active role in eliciting maternal behaviour from their mothers.
In this lecture we
will explore some of the factors that control this reciprocal
partnership. Rat mothers are
attracted by the sight, sound and odour of their pups. Pups utilize
perioral sensations to
locate and suckle from their mother's nipples.
The
lecture concludes with a model put
forward by Carlson that suggests how estrogen interacts with brain
regions involved in
maternal behaviours. You should link this model with the material we
covered on hormonal regulation of
maternal behaviour
Sensory factors in maternal behaviour
|
Compared to people who do not have children, new mothers find the smell of clothes worn by a new-born baby pleasant (Fleming et al, 1993). One of the most surprising abilities of new mothers is their ability to recognize their child on the basis of smell. Kaitz (1987) presented new mothers with a container containing the clothes worn either by their own, or a stranger's baby. This diagram shows the percentage of women who successfully distinguished the odour of their own baby from a stranger's child. Mothers were able to recognize their own baby's smell after only 10 minutes exposure to their baby. |
A virgin female rat does not normally approach a rat pup; in fact when she encounters one, she retreats from the pup as if she was repelled by the pup's odour. However, if virgin rats are continuously exposed to rat pups over several days, they eventually exhibit maternal behaviours and will retrieve pups and adopt the nursing posture even though they do not lactate. This process is called sensitization.
Olfaction plays an important role in inhibiting maternal behaviour in nonpregnant rats.
Mayer & Rosenblatt (1980) confirmed that initial avoidance of pups is based on their odour. They rinsed the olfactory mucosa (lining of the nose) of virgin female rats with zinc sulphate, which temporarily eliminates olfactory sensitivity. Animals that cannot smell are called anosmic , they experience a state called anosmia. Zinc sulphate treatment abolished the virgins' natural aversion to pups, and they started taking care of them. Thus, sensitization involves overcoming a natural aversion to the odour of pups.
| This diagram shows the
initiation of maternal behaviour by nonpregnant (virgin) females
following Hysterectomy+Ovariectomy (HO) + estrogen treatment and/or
olfactory-vomeronasal deafferentation by intranasal zinc sulphate
(ZnSO4) which renders the rats anosmic - unable to smell. Rats were
hysterectomized and ovariectomized in order to remove natural
fluctuating sources of estrogen and progesterone from the body.
Virgin female rats were exposed to groups of 4 pups, 3-8 days of age, during the dark phase of the daily 12:12 hr cycle. Females were considered to have initiated maternal behaviour when they retrieved all test pups during two tests one day apart. Treatment designations:
|
 |
Auditory cues and maternal behaviour
Rat pups emit ultrasonic calls (sounds that we cannot hear) that can be detected via a "bat detector". If the pup gets cold - because it is outside the nest - it emits a characteristic call that allows the mother to locate the pup so that it can be picked up and taken back to the nest. But an experiment by Smotherman et al (1974) shows that rat mothers only respond to this auditory cue if it is accompanied by an olfactory cue from the pup.
 Smotherman's experiment involved giving mothers
who
had delivered a litter of pups, a series of tests in the apparatus
shown here. Stimuli
which differed in their auditory and olfactory
qualities were placed in each
choice arm of the apparatus:
Smotherman's experiment involved giving mothers
who
had delivered a litter of pups, a series of tests in the apparatus
shown here. Stimuli
which differed in their auditory and olfactory
qualities were placed in each
choice arm of the apparatus:
The stimuli were hidden behind a screen at the end of each arm to prevent the female basing her choice on visual cues.
 This
diagram shows the
mothers' preferences when faced with each stimulus combination:
This
diagram shows the
mothers' preferences when faced with each stimulus combination:
mothers showed no preference for:
These results suggest that the smell of pups outside the nest is required before mothers use auditory cues to locate their displaced pups and retrieve them to the nest.
In
other words, mothers only respond to the
auditory cue when it occurs in the presence of pup odour.
Perioral sensation and retrieving
 Tactile
(touch) cues are very important in controlling a number of rat
behaviours.
Tactile
(touch) cues are very important in controlling a number of rat
behaviours.
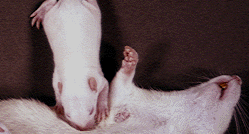
The photograph shows the location of the vibrissae (whiskers) in adult rats and pups.
You can see from this photograph just how delicate the mother's retrieving behaviour must be in order to prevent damaging the pup's skin.
Sensation from the region around the mouth (the perioral area) can be abolished by injecting a local anaesthetic (e.g. lidocaine) into the mystacial (vibrissal) pads which blocks conduction along the infraorbital nerve which conducts tactile information from the whiskers to the brain).
Rats lacking perioral sensation are called anaptic, they are said to be in a state of anapsis.
This
procedure is done under general
anaesthetic in order to minimize any distress experienced by the animal
due to injection.

 This
diagram shows the
results of an experiment (Kenyon et al 1983) that examined the role of
perioral sensation
in maternal behaviour.
This
diagram shows the
results of an experiment (Kenyon et al 1983) that examined the role of
perioral sensation
in maternal behaviour.
Under general anaesthetic lidocaine or saline was injected into
of lactating female rats.
Injecting lidocaine into the vibrissal pads, reduced the number of pups that were retrieved in tests given 30 and 60 minutes after injection. As the effects of the local anaesthetic wore off (120 and 180 minute tests) retrieving behaviour recovered.
Injecting lidocaine into the masseter muscles had no effect on the mothers' ability to retrieve her pups. Thus the effect of lidocaine is site-specific which indicates that lidocaine is not having a general debilitating effect on the animal's behaviour.
| Point to ponder:
Rodents emit ultrasonic call (sounds that we cannot hear) that can be heard via a "bat detector". If the pup gets cold - because it is outside the nest - it emits a characteristic call that triggers the mother into retrieving it back into the nest. Rat pups also emit distress calls if they are roughly handled. What experiment(s) would rule out the possibility that the retrieval deficit produced by lidocaine is a secondary effect mediated by pups' reactions to being roughly treated by lidocaine-injected mothers? |
Perioral
sensation and suckling
|
This has been established by anaesthetizing the mother rat (called a dam) and either
|
 |
This diagram (Kenyon et
al 1982) shows the percentage of pups attaching to the nipples of an
anaesthetized dam after injection of saline or lidocaine into the
vibrissal pads, or into the region of the masseter muscle.
The results indicate that
The effect of lidocaine is site-specific. Injecting local anaesthetic into the pups' masseter muscles does not prevent pups from attaching to their mother's nipples.
|
| In
this experiment Blass et al (1977) found that removing the odour from
the nipples of lactating female rats made them so unattractive that
hardly any rat pups were prepared to attach to them.
However pups were prepared to suckle if the nipples were painted with an extract from the original washings. This result was interpreted as indicating that pups used an olfactory (smell) cue to locate their mother's nipples.
|
 |
A
model of sensory regulation of maternal behaviour in the female rat.
Stern (1989) suggests that there is a complex and integrated 'dance'
between rats pups and
their mothers that 'choreograph' maternal behaviours.
When pups are outside the nest the mother responds to distal cues - the sight, sound and smell of displaced pups. We have seen how the smell of pups is required to motivate the mother to leave her nest, whilst she locates the pups based on their ultrasonic calls.
When the mother comes into contact with her pups proximal cues stimulate her to nuzzle, lick and hover over her pups. We have seen that abolishing perioral sensation by injecting lidocaine into the mother's snout abolishes retrieving behaviour.
Pups also play a more active role in influencing mothers' behaviour. We have seen that abolishing perioral sensation disrupts pups' ability to attach to their mother's nipples. But in addition, when pups nuzzle their mother's ventral surface (belly) the mother becomes still, her back arches and she crouches over her pups. Stern calls this behaviour kyphosis and clearly it would aid pups find and attach to nipples.
Stern (1989) reviews a number of experiments which show that kyphosis is disrupted
This
table summarizes the parts played by
sensory factors in the interactions between mother rats and their
offspring.
| Mother |
Pups |
 |
|
| Distal cues stimulate mother to seek contact with pups | Pups provide distal cues: sight, sound, and smell for the mother |
| Perioral contact with her pups stimulates the mother to nuzzle, lick and hover over pups | The mother makes perioral contact with pups |
| Pups making ventral contact with the mother stimulates her to adopt the crouching posture | The pups nuzzle the mother's ventral surface. The crouching posture facilitates the pups' suckling behaviour |
Neural
factors in maternal behaviour
The
medial preoptic area
So far, this lecture has focussed on the role of sensory factors in maternal behaviour. For example, you now appreciate that anosmia facilitates the onset of maternal behaviour, and that intact perioral sensation is a prerequisite for retrieval.
In a previous lecture we saw that
Quite clearly the brain must play an important role in mediating the effects of hormones and sensory factors on maternal behaviour.
Where in the brain does this integration take place? One possibility is that estrogen overcomes a rat's natural aversion to the smell of pups, and that this process occurs in an area of the hypothalamus called the medial preoptic area (MPOA).
There are several reasons for focussing on the MPOA:
Numan et al (1977) investigated the possibility that the MPOA was the area of the brain where estrogen influences maternal behaviour. In this experiment all the subjects were virgin rats who were mated and on Day 16 of pregnancy they were all hysterectomized and ovariectomized. You will recall from a previous lecture that maternal behaviour is facilitated by the rise in estrogen which occurs shortly after pregnant rats are hysterectomised on Day 16 of pregnancy. But this facilitation of maternal behaviour does not occur if the rats are also ovariectomised. Ovariectomy removes a natural source of estrogen. Thus the 16 Day hysterectomised-ovariectomized pregnant rat can be used to study the role of estrogen in the onset of maternal behaviour.
Numan et al (1977) studied five independent groups of rats that had estrogen injections in various areas on Day 16 of pregnancy:
Cholesterol was used as a placebo control. Cholesterol is a steroid which has a similar chemical structure to estradiol. Implanting estrogen into the VMH, MB and subcutaneously was done to check that any effects produced by MPOA implants were site-specific.
The diagram below shows the cumulative percentage of 16 day pregnant hysterectomised (H) and ovariectomised (O) female rats showing maternal behaviour over the five day test period (Numan et al 1977). Compared to the other groups, rats with estradiol implanted into the MPOA show rapid onset of maternal behaviours when they are presented with foster pups. You can use this diagram to satisfy yourself that the MPOA is the most effective site for implanting estrogen to facilitate maternal behaviour.
Carlson and
Numan's model of hormone effects on maternal behaviour
 This
diagram that shows how various
interconnected areas of the brain could be involved in the rat's
reaction to pup odour.
This
diagram that shows how various
interconnected areas of the brain could be involved in the rat's
reaction to pup odour.
Olfactory information about the pup is received by the olfactory bulbs, then passed to the medial nuclei of the amygdala via the lateral olfactory tract.
The amygdala is thought to exert an inhibitory influence over the medial preoptic area (MPOA) which is transmitted down the stria terminalis
The output of the MPOA passes through the ventral tegmental area to terminate in areas of the brain involved in motor functions.
Carlson
(2001) reviews the evidence that
supports this model.
 |
Here
are the components of Numan and Carlson's model highlighted on a
sagittal section of the rat brain.
Lesioning:
Lesioning:
Medial amygdala inhibition of the MPOA can be overcome by injecting estradiol directly into the MPOA. |
The ventral tegmental area (VTA)
In the diagram of Carlson's model the MPOA is depicted as an area of the brain which receives hormonal (estrogen) and sensory (odour) information, integrates this information, and sends instructions to motor areas of the brain to perform maternal behaviour. The nerves that carry these instructions pass through the VTA. Consequently VTA lesions disrupt behaviour because they destroy this pathway which is responsible for the initiation of motor activity.
But Stern (1989) has argued that the effects of VTA lesions can be explained in a completely different way. She points out that the nerves carrying sensory information from the rat's snout run across the top of VTA. This pathway is called the trigeminal sensory pathway. Stern argues that VTA lesions would also damage this sensory pathway. She argues that damage to this pathway is responsible for the effects seen in rats with VTA lesions.
Earlier we studied an experiment in which retrieving behaviour was abolished after lidocaine was injected into the snout to block conduction in the trigeminal nerve. Stern suggests that sensory information travelling through the trigeminal pathway may reach the MPOA. She argues that the reason MPOA land VTA lesions disrupt maternal behaviour is because the mother rat is unable to process the sensory tactile information from pups that controls her behaviour.
In a nutshell:
| Click
the links to explore:
|
We have already mentioned that electrical lesions which destroy the VTA - and any nerve fibres passing through the VTA - severely disrupt maternal behaviour. This result is predicted by both theories. Furthermore, maternal behaviour is not disrupted if a chemical - that destroys cells in the VTA, but does not interfere with nerve fibres that travel through the VTA - is injected into the VTA. Again this result would be predicted by both theories. You can explore the effects of chemical and electrical lesions and the two theories in the diagram above.
Testing Stern's and Numan's theories
One way of testing Numan and Stern's theories would be to compare the effects outside the VTA on maternal behaviour of lesions :
 Numan
and Numan (1991)
report that "coronal knife cuts located in the midbrain tegmentum at a
point caudal
to the VTA severely disrupted maternal behaviour", but perioral tactile
sensitivity
was not affected by this operation.
Numan
and Numan (1991)
report that "coronal knife cuts located in the midbrain tegmentum at a
point caudal
to the VTA severely disrupted maternal behaviour", but perioral tactile
sensitivity
was not affected by this operation.
These results support their suggestion that a pathway running from the MPOA through the VTA projects to brain areas involved in motor functions.
It is difficult to explain these results in terms of the lesion interfering with perioral sensation, because a test of perioral sensation showed that there was no difference between sham operated and lesioned rats.
One way to test Stern's theory is to cut the infraorbital branch of the trigeminal nerve that carries sensory information from the rat's snout to its brain.
 |
Kenyon
et al (1983) cut the infraorbital branch of the trigeminal nerves which
transmitting sensory information from the mystacial pads to the brain.
The results shown here indicate a profound but short-lasting disruption of maternal behaviour after this operation. In addition perioral tactile sensation was abolished by this operation. Kenyon and Numan's results indicate that maternal behaviour is disrupted by:
|
Online text resources:
 |
|
Supplementary
material
 There are some impressive sets of vibrissae out there! As you may have
realised during the lecture I am very interested in the role played by
vibrissae in controlling various aspects of rat behaviour. Here are
some sources of further information. It is worth looking at some of
these links to get a feel for how study of an apparently esoteric
sensory system in the rat may lead to fundamental breakthroughs in our
understanding of the relationship between brain and behaviour.
There are some impressive sets of vibrissae out there! As you may have
realised during the lecture I am very interested in the role played by
vibrissae in controlling various aspects of rat behaviour. Here are
some sources of further information. It is worth looking at some of
these links to get a feel for how study of an apparently esoteric
sensory system in the rat may lead to fundamental breakthroughs in our
understanding of the relationship between brain and behaviour.
|

 The
sense
of smell (olfaction) plays an important
role in the maternal behaviour of humans and other animals.
The
sense
of smell (olfaction) plays an important
role in the maternal behaviour of humans and other animals.